
Estuvimos casi media hora pensando en cómo partir este post sin parecer vulgares (o sin parecer que somos parte de la Asexma). Porque es un tema sin duda interesante, pero que hará sonrojar a más de un conservador. Pero la pregunta es simple: ¿qué pasó con nuestro hueso peneano? ¿Lo teníamos? En el caso del «negro de Whatsapp», ¿es un fémur?
Lo siento, no pudimos (N.d.E.: a pesar de que se lo advertimos)
Como usted ya debe saber, efectivamente el pene en los machos humanos no tiene hueso (es verdad, no insista). Sin embargo, algunos de nuestros parientes cercanos, como macacos y bonobos, sí lo tienen, aunque sea pequeño. Recordemos que éstos pasan gran parte del día teniendo todo tipo de relaciones sexuales. ¿Podría haber alguna relación?
Este simpático y juguetón huesito es conocido en el mundo de las ciencias como «báculo», «baculum» o simplemente «hueso peneano».
Pero vamos con algunos datos interesantes sobre el báculo (1313). Este hueso es considerado como «el más diverso de todos los huesos» [1], ya que presenta una enorme variedad de tamaños, formas, ancho, largo, etc. en aquellos mamíferos que lo poseen [2].

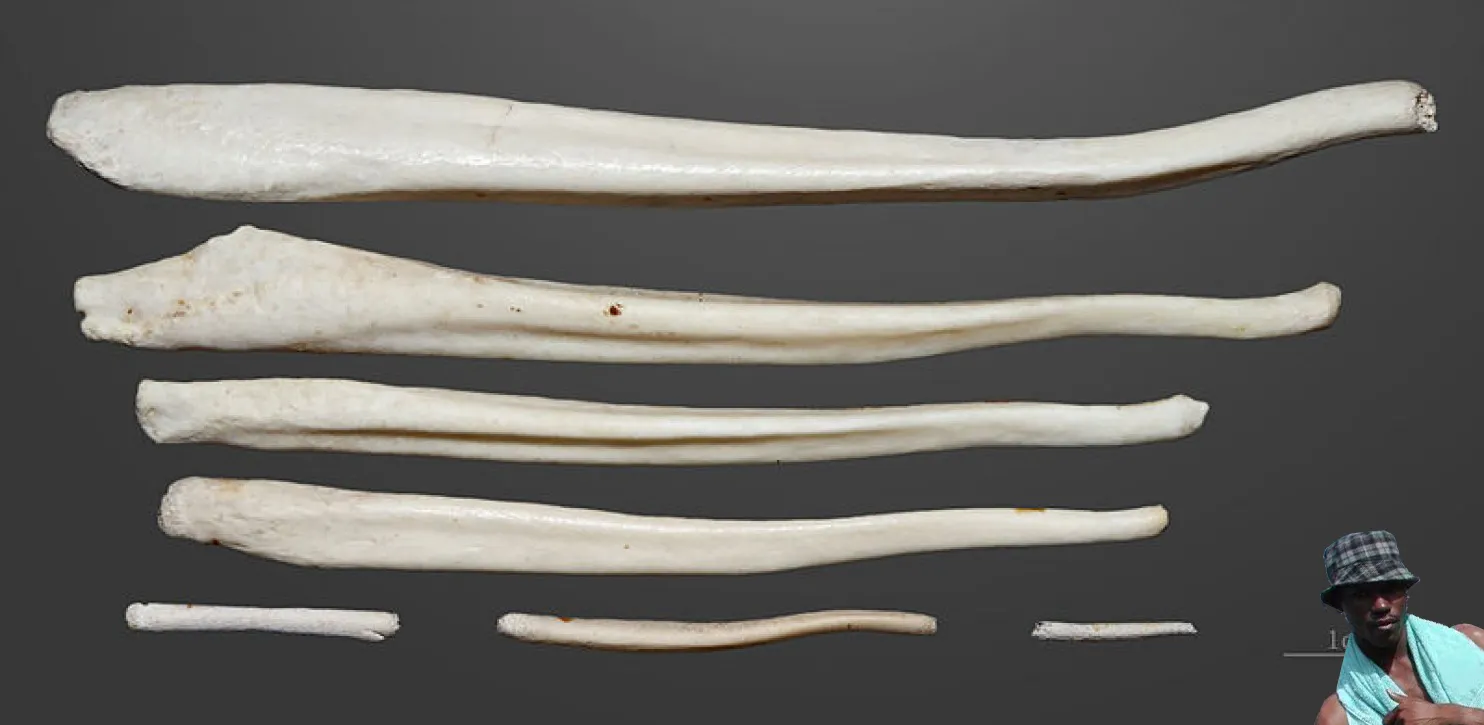
Baculum de un macaco japonés o de cara roja:Macaca fuscata . No se nos ocurrió ningún chiste.
Algo relevante al respecto es que el báculo no se encuentra de forma uniforme entre los mamíferos. De hecho, sólo es posible encontrarlo en 8 órdenes: Afrosoricida (tenrecs y topos dorados), Carnivora (osos, cánidos, felinos, etc), Chiroptera (murciélagos), Dermoptera (lemures voladores), Erinaceomorpha (erizos de tierra), Primates (gorilas, orangutanes, bonobos y quienes escriben), Rodentia (ratones, hamsters, etc.) y Soricomporpha (musarañas, tálpidos, solenodones, etc.) [3,4]. Recientemente se ha descubierto a un lagomorfo (como los conejos) que también posee un pequeño báculo [5], a pesar de que en este orden no había sido descrito previamente. Esto sugiere que la presencia de este hueso es mucho más prevalente entre los mamíferos de lo que pensábamos.
Como usted puede imaginar, dentro de todos estos órdenes en donde se ha descrito la presencia del báculo hay, por cierto, excepciones. Por ejemplo, dentro de los primates, los tarsios (Tarsiidae), algunos monos del nuevo mundo (Platyrrhines) y, por supuesto, nosotros los humanos, carecemos de tal hueso.
Una de las hipótesis que se han propuesto para las grandes variaciones morfológicas en el báculo entre los mamíferos es que esta se debe al resultado de la selección sexual (particularmente post-copulatoria). La morfología genital está sin duda expuesta a este tipo de presiones selectivas.

Cuando hay mucha competencia, todo vale.
Hay algunos estudios que sugieren que el tamaño del báculo en algunos roedores poligámicos (Mus domesticus) es un predictor significativo del éxito reproductivo en machos [6]. Otros estudios incluso han logrado manipular de forma experimental el báculo, alterando las presiones de selección sexual en ratones. Es decir, ofreciendo a un grupo de ratones muchas hembras y otros una única hembra en ausencia de otros machos. Durante los experimentos, mientras el grupo experimental estaba luchando arduamente por poder traspasar su material genético, compitiendo con varios machos igual de capaces, sus amigos del grupo de control ya se estaban fumando un cigarro y roncando a pata suelta.
Luego de 27 generaciones, aquellas poblaciones con niveles forzosamente altos de competencia post-copulatoria (altas presiones de selección sexual), desarrollaban báculos mucho más gruesos que sus amigos a los cuales los habían forzado a la monogamia (donde la selección sexual está ausente). ¡Mirá de quién te burlaste!
¿Qué nos hace pensar esto?
Bueno, que las presiones de selección de tipo sexual podrían estar modelando no sólo los tamaños y formas, sino la presencia/ausencia de este hueso.
Basado en esta idea, y para intentar establecer alguna hipótesis acerca de por qué algunos primates tienen báculo y otros no, es en donde se encuentra emplazado el estudio recientemente publicado que hoy nos convoca: ¿tendrán que ver nuestras estrategias copulatorias con la ausencia de báculo en los humanos?
Brindle y Opie [8] intentaron «trazar» la historia evolutiva de este hueso a través del tiempo para poder reconstruir algunos estados ancestrales y tratar de establecer dónde y bajo qué condiciones perdimos esta característica. Algo importante, y aquí nos ponemos a hablar en jerigoncio, es que los autores quieren poner a prueba que la «hipótesis de intromisión (if you know what I mean ;) ) prolongada», o para los amigos, qué tanto tiempo penetra el pene durante la cópula, sería un predictor para la presencia/ausencia del báculo a lo largo de la evolución de los primates. Esto se basa en que la presencia de este hueso facilita intromisiones prolongadas e incrementa la proximidad al cervix, lo que reduce los niveles de competencia espermática y, por lo tanto, aumenta el éxito reproductivo.
¿Dónde hay más competencia espermática? Bueno, justamente en especies poligámicas, donde muchos machos copulan con una misma hembra y todos esos espermatozoides de distintos machos deben competir por fecundar. Los autores señalan que «la duración de la intromisión se espera que esté relacionada con la presencia del báculo en el curso de la evolución. Especies con intromisiones prolongadas deberían tener báculos elongados. Y, por último, aquellos grupos en donde las presiones selectivas de tipo sexual, particularmente las post-copulatorias son altas, como aquellas especies poligámicas, deberían tener báculos más grandes que aquellos con niveles más bajos de presiones post-copulatorias».
Nos encantaría darle detalles de cómo lo hicieron, pero mi tío Markov es un poco tímido. Iremos directo a lo que nos interesa.
El principal resultado es que el baculum evolucionó primero entre 145 y 95 millones de años atrás, según los autores, lo que sugiere que estaba presente en el ancestro común de todos los primates y carnívoros [8]. Una de las posibles razones que proponen los autores para la pérdida del huesito en humanos tiene que ver justamente con las distintas prácticas reproductivas: en primates, la presencia del hueso peneano estaría relacionada con el incremento en la duración de la intromisión. De hecho, los autores muestran evidencia positiva significativa para una correlación entre la presencia del báculo y la duración de la intromisión en primates [8].
En este sentido, intromisiones más largas a menudo ocurren en especies con prácticas poligámicas, como en bonobos y chimpancés, pero no en humanos. Esta estrategia implica una competencia intensa por la cópula (fecundación) y una forma de limitar el acceso de otros machos a las hembras es, justamente, acaparar: pasar más tiempo copulando con una misma hembra para que esta no esté disponible, aumentando la probabilidad de éxito de la fecundación en un ambiente competitivo. Este hueso facilita esta estrategia, apoyando el pene durante el sexo, manteniendo la uretra abierta y evitando cualquier constricción.
En humanos esto no ocurre. De hecho, hay machos que gastan muy poco tiempo en la intromisión :( (N.d.E.: =( ). Pero, si nos abstraemos de aspectos culturales del sexo y pensamos que el objetivo del varón es sólo eyacular dentro de la hembra para poder fecundarla y así transferir su material genético (los científicos somos muy preco... fríos... digo), podríamos sugerir que los humanos tenemos tiempos de intromisión muy reducidos. Como los humanos tendemos a ser monógamos (o más bien la poligamia no es tan común), la competencia post-copulatoria es muy baja, debido a que, en general, la hembra copula con un solo macho (ahí hay otro tipo de competencia, basada en qué espermatozoide será finalmente el que fecunde). De hecho, los autores señalan que el hecho de que no tengamos intromisiones muy prolongadas ni competencia post-copulatoria «nos podría dar luces acerca de por qué el báculo se perdió en el linaje de los humanos».
Ahora, usted se preguntará: «pero si no tenemos hueso, ¿por qué se fractura?».
Y es cierto: esto ocurre en humanos machos. Pero, más que una fractura, se trata de un desgarro de los cuerpos cavernosos.
Al lector macho lo dejo con esa imagen mental. La lectora hembra, si lo desea, puede obviar ese cuadro (o reírse de él).
Referencias
1.- Patterson BD, Thaeler CS. 1982 The mammalian baculum: hypotheses on the nature of bacular variability. J. Mammal. 63, 1–15. (doi:10.1890)
2.- Eberhard W. 1993 Evaluating models of sexual selection: genitalia as a test case. Am. Nat. 142, 564– 571. (doi:10.1086/285556)
3.- Perrin WF, Wursig B, Thewissen JGM (eds). 2009 Encyclopaedia of marine mammals, 2nd edn. London, UK: Academic Press.
4.-Martin RD. 2007 The evolution of human reproduction: a primatological perspective. Yearb. Phys. Anthropol. 50, 59 – 84. (doi:10.1002/ ajpa.20734)
6.- Stockley P, Ramm SA, Sherborne AL, Thom MDF, Paterson S, Hurst JL. 2013 Baculum morphology predicts reproductive success of male house mice under sexual selection. BMC Biol. 11, 66. (doi:10. 1186/1741-7007-11-66)
7.- Simmons LW, Firman RC. 2013 Experimental evidence for the evolution of the mammalian baculum by sexual selection. Evolution 68, 276 – 283. (doi:10.1111/evo.12229) 9. Harcourt AH, Harvey PH, Larson SG, Sh
8.- Matilda Brindle and Christopher Opie, 2016. Postcopulatory sexual selection influences baculum evolution in primates and carnivores. Proceeding of the Royal Society B. 283: 20161736.
